Ekspert medyczny artykułu

Nowe publikacje
Jajnik
Ostatnia recenzja: 04.07.2025

Cała zawartość iLive jest sprawdzana medycznie lub sprawdzana pod względem faktycznym, aby zapewnić jak największą dokładność faktyczną.
Mamy ścisłe wytyczne dotyczące pozyskiwania i tylko linki do renomowanych serwisów medialnych, akademickich instytucji badawczych i, o ile to możliwe, recenzowanych badań medycznych. Zauważ, że liczby w nawiasach ([1], [2] itd.) Są linkami do tych badań, które można kliknąć.
Jeśli uważasz, że któraś z naszych treści jest niedokładna, nieaktualna lub w inny sposób wątpliwa, wybierz ją i naciśnij Ctrl + Enter.
Jajnik (ovarium; gr. oophoron) jest parzystym narządem, żeńskim gruczołem rozrodczym, zlokalizowanym w jamie miednicy za szerokim więzadłem macicy. W jajnikach rozwijają się i dojrzewają żeńskie komórki rozrodcze (jaja), a także powstają żeńskie hormony płciowe, które przedostają się do krwi i limfy. Jajnik ma jajowaty kształt, nieco spłaszczony w kierunku przednio-tylnym. Kolor jajnika jest różowawy. Na powierzchni jajnika kobiety, która urodziła, widoczne są zagłębienia i blizny - ślady owulacji i przekształcenia ciałek żółtych. Jajnik waży 5-8 g. Wymiary jajnika wynoszą: długość 2,5-5,5 cm, szerokość 1,5-3,0 cm, grubość - do 2 cm. Jajnik ma dwie wolne powierzchnie: powierzchnię przyśrodkową (facies medialis), zwróconą do jamy miednicy, częściowo przykrytą jajowodem, oraz powierzchnię boczną (facies lateralis), przylegającą do ściany bocznej miednicy, do lekko zaznaczonego zagłębienia - dołu jajnikowego. Dół ten znajduje się w kącie między naczyniami biodrowymi zewnętrznymi pokrytymi otrzewną u góry a tętnicami macicznymi i zasłonowymi u dołu. Za jajnikiem moczowód odpowiedniej strony przebiega retroperitonealnie z góry na dół.
Powierzchnie jajnika przechodzą w wypukły wolny (tylny) brzeg (margo liber), z przodu - w brzeg krezkowy (margo mesovaricus), przyczepiony za pomocą krótkiego fałdu otrzewnej (krezki jajnika) do tylnego płatka szerokiego więzadła macicy. Na tym przednim brzegu narządu znajduje się wyżłobione zagłębienie - wnęka jajnika (hilum ovarii), przez którą tętnica i nerwy wchodzą do jajnika, a żyły i naczynia limfatyczne wychodzą. Jajnik ma również dwa końce: zaokrąglony górny koniec rurkowaty (extremitas tubaria), zwrócony w stronę jajowodu, oraz dolny koniec maciczny (extremitas utenna), połączony z macicą za pomocą więzadła jajnikowego (lig. ovarii proprium). To więzadło, w postaci okrągłego sznura o grubości około 6 mm, biegnie od końca macicznego jajnika do kąta bocznego macicy, znajdującego się między dwoma liśćmi więzadła szerokiego macicy. Aparat więzadłowy jajnika obejmuje również więzadło wieszadłowe jajnika (lig.suspensorium ovarii), które jest fałdem otrzewnej, który biegnie od ściany miednicy małej do jajnika i zawiera wewnątrz naczynia jajnikowe i wiązki włókien włóknistych. Jajnik jest unieruchomiony przez krótką krezkę (mesovarium), która jest duplikacją otrzewnej, która biegnie od tylnego płatka więzadła szerokiego macicy do krawędzi krezkowej jajnika. Same jajniki nie są pokryte otrzewną. Największy frędzel jajnikowy jajowodu jest przyczepiony do rurkowatego końca jajnika. Topografia jajnika zależy od położenia macicy, jej wielkości (podczas ciąży). Jajniki są bardzo ruchomymi narządami małej miednicy.
 [
[ Naczynia i nerwy jajnika
Dopływ krwi do jajników zapewniają aa. et vv. ovaricae et uterinae. Obie tętnice jajnikowe (aa. ovaricae dextra et sinistra) biorą swój początek na przedniej powierzchni aorty tuż pod tętnicami nerkowymi; prawa zwykle bierze początek z aorty, a lewa z tętnicy nerkowej. Kierując się w dół i bocznie wzdłuż przedniej powierzchni mięśnia biodrowo-lędźwiowego większego, każda tętnica jajnikowa przecina moczowód z przodu (oddając mu gałęzie), naczynia biodrowe zewnętrzne, linię graniczną i wchodzi do jamy miednicy, znajdując się tutaj w więzadle wieszadłowym jajnika. Podążając w kierunku przyśrodkowym, tętnica jajnikowa przechodzi między liśćmi więzadła szerokiego macicy pod jajowodem, oddając mu gałęzie, a następnie do krezki jajnika; wchodzi do wnęki jajnika.
Gałęzie tętnicy jajnikowej szeroko zespalają się z gałęziami jajnikowymi tętnicy macicznej. Odpływ żylny z jajników odbywa się przede wszystkim do splotu żylnego jajnika, zlokalizowanego w okolicy wnęki jajnika. Stąd odpływ krwi przebiega w dwóch kierunkach: przez żyły maciczne i jajnikowe. Prawa żyła jajnikowa ma zastawki i uchodzi do żyły głównej dolnej. Lewa żyła jajnikowa uchodzi do lewej żyły nerkowej, chociaż nie ma zastawek.
Odpływ limfatyczny z jajników odbywa się przez naczynia limfatyczne, szczególnie obficie w okolicy wrót narządowych, gdzie wyróżnia się splot limfatyczny podjajnikowy. Następnie limfa odprowadzana jest wzdłuż naczyń limfatycznych jajników do węzłów chłonnych przyaortalnych.
Unerwienie jajników
Układ współczulny - zaopatrywany przez włókna pozwojowe ze splotu trzewnego (słonecznego), krezkowego górnego i podbrzusznego; układ przywspółczulny - za pośrednictwem nerwów trzewnych krzyżowych.
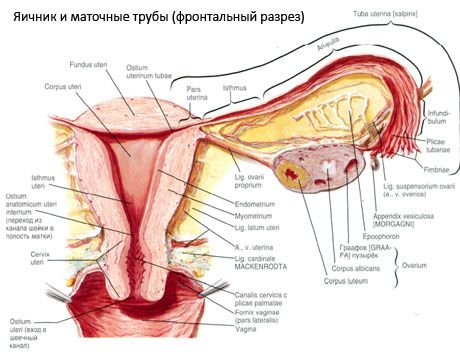
Budowa jajnika
Powierzchnia jajnika pokryta jest jednowarstwowym nabłonkiem zarodkowym. Pod nim znajduje się gęsta błona białkowa tkanki łącznej (tunica albuginea). Tkanka łączna jajnika tworzy jego podścielisko (stroma ovarii), bogate we włókna sprężyste. Substancja jajnika, jego miąższ, dzieli się na warstwę zewnętrzną i wewnętrzną. Warstwa wewnętrzna, znajdująca się w centrum jajnika, bliżej jego wrót, nazywana jest rdzeniem (medulla ovarii). W tej warstwie, w luźnej tkance łącznej, znajdują się liczne naczynia krwionośne i limfatyczne oraz nerwy. Gęstsza jest warstwa zewnętrzna jajnika - kora (cortex ovarii). Zawiera dużo tkanki łącznej, w której zlokalizowane są dojrzewające pęcherzyki jajnikowe pierwotne (folliculi ovarici primarii), pęcherzyki wtórne (pęcherzykowe) (folliculi ovarici secundarii, s.vesiculosi), a także pęcherzyki dojrzałe, pęcherzyki Graafa (folliculi ovarici maturis) oraz ciałka żółte i atretyczne.
Każdy pęcherzyk zawiera żeńską komórkę rozrodczą, czyli oocyt (ovocytus). Komórka jajowa ma średnicę do 150 µm, jest okrągła, zawiera jądro, dużą ilość cytoplazmy, która oprócz organelli komórkowych zawiera wtrącenia białkowo-lipidowe (żółtko), glikogen, niezbędny do odżywiania komórki jajowej. Komórka jajowa zazwyczaj zużywa zapas składników odżywczych w ciągu 12-24 godzin po owulacji. Jeśli nie dojdzie do zapłodnienia, komórka jajowa obumiera.
Ludzkie jajo ma dwie błony, które je pokrywają. Wewnątrz znajduje się cytolemma, czyli błona cytoplazmatyczna jaja. Na zewnątrz cytolemma znajduje się warstwa tzw. komórek pęcherzykowych, które chronią jajo i pełnią funkcję hormonotwórczą - wydzielają estrogeny.
Fizjologiczną pozycję macicy, jajowodów i jajników zapewniają aparaty podtrzymujące, mocujące i podtrzymujące, które łączą otrzewną, więzadła i tkankę miednicy. Aparat podtrzymujący jest reprezentowany przez formacje parzyste, obejmuje więzadła okrągłe i szerokie macicy, więzadła właściwe i więzadła podtrzymujące jajników. Więzadła szerokie macicy, więzadła właściwe i podtrzymujące jajników utrzymują macicę w położeniu środkowym. Więzadła okrągłe ciągną dno macicy do przodu i zapewniają jej fizjologiczne pochylenie.
Aparat mocujący (kotwiczący) zapewnia położenie macicy w centrum miednicy małej i praktycznie uniemożliwia jej przesuwanie się na boki, do przodu i do tyłu. Ponieważ jednak aparat więzadłowy odchodzi od macicy w jej dolnej części, możliwe są przechyły macicy w różnych kierunkach. Aparat mocujący obejmuje więzadła zlokalizowane w luźnej tkance miednicy i rozciągające się od dolnej części macicy do bocznych, przednich i tylnych ścian miednicy: więzadła sakralne, kardynalne, maciczno-pęcherzowe i pęcherzowo-łonowe.
Oprócz krezki jajnikowej wyróżnia się następujące więzadła jajnikowe:
- więzadło wieszadłowe jajnika, wcześniej nazywane więzadłem lejkowo-miedniczkowym. Jest to fałd otrzewnej, przez który przechodzą naczynia krwionośne (a. et v. ovarica) oraz naczynia limfatyczne i nerwy jajnika, rozciągnięty między boczną ścianą miednicy, powięzią lędźwiową (w rejonie podziału tętnicy biodrowej wspólnej na zewnętrzną i wewnętrzną) i górnym (jajowodowym) końcem jajnika;
- właściwe więzadło jajnika przechodzi między listkami szerokiego więzadła macicy, bliżej tylnego liścia, i łączy dolny koniec jajnika z boczną krawędzią macicy. Właściwe więzadło jajnika jest przyczepione do macicy między początkiem jajowodu a więzadłem obłym, za i nad tym ostatnim. W grubości więzadła przechodzą rr. ovarii, które są końcowymi gałęziami tętnicy macicznej;
- więzadło wyrostkowo-jajnikowe Więzadło rozciąga się od krezki wyrostka robaczkowego do prawego jajnika lub więzadła szerokiego macicy w postaci fałdu otrzewnej. Więzadło jest niestałe i występuje u 1/2 - 1/3 kobiet.
Aparat podporowy stanowią mięśnie i powięzie dna miednicy, podzielone na warstwę dolną, środkową i górną (wewnętrzną).
Najsilniejsza jest górna (wewnętrzna) warstwa mięśni, reprezentowana przez sparowany mięsień, który podnosi odbyt. Składa się z wiązek mięśni, które rozchodzą się wachlarzowato od kości guzicznej do kości miednicy w trzech kierunkach (łonowo-guziczny, biodrowo-guziczny i kulszowo-goleniowy). Ta warstwa mięśni jest również nazywana przeponą miednicy.
Środkowa warstwa mięśni znajduje się między spojeniem łonowym, kością łonową i kulszową. Środkowa warstwa mięśni - przepona moczowo-płciowa - zajmuje przednią połowę ujścia miednicy, przez którą przechodzi cewka moczowa i pochwa. W przedniej części między jej płatami znajdują się wiązki mięśni, które tworzą zwieracz zewnętrzny cewki moczowej, w tylnej części znajdują się wiązki mięśni, które biegną w kierunku poprzecznym - głęboki mięsień poprzeczny krocza.
Dolna (zewnętrzna) warstwa mięśni dna miednicy składa się z mięśni powierzchownych, które kształtem przypominają cyfrę 8. Należą do nich mięsień opuszkowo-jamisty, mięsień kulszowo-jamisty, zwieracz zewnętrzny odbytu i powierzchowny mięsień poprzeczny krocza.
Ontogeneza jajników
Proces wzrostu pęcherzyków i atrezji rozpoczyna się w 20. tygodniu ciąży, a do momentu porodu w jajnikach dziewczynki pozostaje do 2 milionów oocytów. Do pierwszej miesiączki ich liczba spada do 300 tysięcy. W całym okresie życia rozrodczego nie więcej niż 500 pęcherzyków osiąga dojrzałość i owuluje. Początkowy wzrost pęcherzyków nie zależy od stymulacji FSH, jest ograniczony, a atrezja szybko występuje. Uważa się, że zamiast hormonów steroidowych głównym regulatorem wzrostu i atrezji pęcherzyków pierwotnych są lokalne peptydy autokrynne/parakrynne. Uważa się, że proces wzrostu pęcherzyków i atrezji nie jest przerywany przez żadne procesy fizjologiczne. Proces ten trwa w każdym wieku, w tym w okresie wewnątrzmacicznym i menopauzie, i jest przerywany przez ciążę, owulację i anowulację. Mechanizm, który wyzwala wzrost pęcherzyków i ich liczba w każdym konkretnym cyklu, jest nadal niejasny.
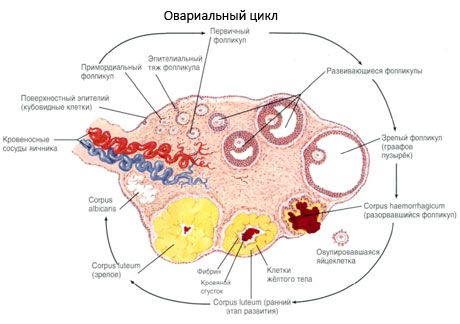
W trakcie rozwoju pęcherzyk przechodzi kilka etapów rozwoju. Pierwotne komórki rozrodcze pochodzą z endodermy woreczka żółtkowego, omoczni i migrują do obszaru genitalnego zarodka w 5-6 tygodniu ciąży. W wyniku szybkiego podziału mitotycznego, który trwa od 6-8 tygodnia do 16-20 tygodnia ciąży, w jajnikach zarodka powstaje do 6-7 milionów oocytów, otoczonych cienką warstwą komórek ziarnistych.
Pęcherzyk preantralny - oocyt otoczony jest błoną (Zona pellucida). Komórki ziarniste otaczające oocyt zaczynają proliferować, ich wzrost zależy od gonadotropin i koreluje z poziomem estrogenów. Komórki ziarniste są celem dla FSH. W fazie pęcherzyka preantralnego komórki ziarniste są zdolne do syntezy trzech klas steroidów: indukuje przede wszystkim aktywność aromatazy, głównego enzymu, który przekształca androgeny w estradiol. Uważa się, że estradiol jest zdolny do zwiększania liczby własnych receptorów, wywierając bezpośredni mitogenny wpływ na komórki ziarniste niezależnie od FSH. Uważa się go za czynnik parakrynowy, który wzmacnia działanie FSH, w tym aktywację procesów aromatyzacji.
Receptory FSH pojawiają się na błonach komórek ziarnistych, gdy tylko rozpoczyna się wzrost pęcherzyka. Spadek lub wzrost FSH prowadzi do zmiany liczby jego receptorów. To działanie FSH jest modulowane przez czynniki wzrostu. FSH działa poprzez białko G, układ cyklazy adenylowej, chociaż steroidogeneza w pęcherzyku jest regulowana głównie przez FSH, w tym procesie bierze udział wiele czynników: kanały jonowe, receptory kinazy tyrozynowej, układ fosfolipazy drugich przekaźników.
Rola androgenów we wczesnym rozwoju pęcherzyka jest złożona. Komórki ziarniste mają receptory androgenowe. Są one nie tylko substratem dla aromatyzacji do estrogenów indukowanej przez FSH, ale mogą również zwiększać aromatyzację przy niskich stężeniach. Gdy poziom androgenów wzrasta, komórki ziarniste preantralne preferencyjnie wybierają nie ścieżkę aromatyzacji do estrogenów, ale prostszą ścieżkę androgenową za pośrednictwem 5a-reduktazy, aby przekształcić się w androgen, który nie może zostać przekształcony w estrogen, tym samym hamując aktywność aromatazy. Ten proces hamuje również powstawanie receptorów FSH i LH, zatrzymując w ten sposób rozwój pęcherzyka.
Proces aromatyzacji, pęcherzyk z wysokim poziomem androgenów ulega procesom atrezji. Wzrost i rozwój pęcherzyka zależy od jego zdolności do przekształcania androgenów w estrogeny.
W obecności FSH dominującą substancją płynu pęcherzykowego będą estrogeny. W przypadku braku FSH - androgeny. LH jest normalnie nieobecny w płynie pęcherzykowym do połowy cyklu. Gdy tylko poziom LH wzrasta, aktywność mitotyczna komórek ziarnistych spada, pojawiają się zmiany zwyrodnieniowe i poziom androgenów w pęcherzyku wzrasta. Poziom steroidów w płynie pęcherzykowym jest wyższy niż w osoczu i odzwierciedla aktywność funkcjonalną komórek jajnika: komórek ziarnistych i tekalnych. Jeśli jedynym celem dla FSH są komórki ziarniste, to LH ma wiele celów - komórki tekalne, komórki podścieliska i luteinowe oraz komórki ziarniste. Zarówno komórki ziarniste, jak i tekalne mają zdolność do steroidogenezy, ale aktywność aromatazy dominuje w komórkach ziarnistych.
W odpowiedzi na LH komórki tekalne produkują androgeny, które następnie są przekształcane przez komórki ziarniste w estrogeny poprzez aromatyzację indukowaną przez FSH.
W miarę wzrostu pęcherzyka komórki tekalne zaczynają wyrażać geny dla receptorów LH, P450 sec i 3beta-hydroksysteroidowej dehydrogenazy, a insulinopodobny czynnik wzrostu (IGF-1) działa synergicznie z LH, zwiększając ekspresję genów, ale nie stymulując steroidogenezy.
Steroidogeneza jajników jest zawsze zależna od LH. W miarę wzrostu pęcherzyka komórki tekalne ekspresują enzym P450c17, który tworzy androgen z cholesterolu. Komórki ziarniste nie mają tego enzymu i są zależne od komórek tekalnych w produkcji estrogenów z androgenów. W przeciwieństwie do steroidogenezy, folikulogeneza jest zależna od FSH. W miarę wzrostu pęcherzyka i wzrostu poziomu estrogenu aktywowany jest mechanizm sprzężenia zwrotnego - produkcja FSH jest hamowana, co z kolei prowadzi do zmniejszenia aktywności aromatazy pęcherzyka i ostatecznie do atrezji pęcherzyka poprzez apoptozę (programowaną śmierć komórki).
Mechanizm sprzężenia zwrotnego estrogenów i FSH hamuje rozwój pęcherzyków, które zaczęły rosnąć, ale nie pęcherzyka dominującego. Pęcherzyk dominujący zawiera więcej receptorów FSH, które wspomagają proliferację komórek ziarnistych i aromatyzację androgenów do estrogenów. Ponadto szlaki parakrynowe i autokrynowe działają jako ważny koordynator rozwoju pęcherzyka antralnego.
Autokrynny/parakrynny regulator składa się z peptydów (inhibiny, aktywiny, folistatyny), które są syntetyzowane przez komórki ziarniste w odpowiedzi na FSH i przedostają się do płynu pęcherzykowego. Inhibina zmniejsza wydzielanie FSH; aktywina stymuluje uwalnianie FSH z przysadki mózgowej i wzmacnia działanie FSH w jajniku; folistatyna hamuje aktywność FSH, prawdopodobnie poprzez wiązanie aktywiny. Po owulacji i rozwoju ciałka żółtego inhibina jest pod kontrolą LH.
Na wzrost i różnicowanie komórek jajnika wpływają czynniki wzrostu podobne do insuliny (IGE). IGF-1 działa na komórki ziarniste, powodując wzrost cyklicznego monofosforanu adenozyny (cAMP), progesteronu, oksytocyny, proteoglikanu i inhibiny.
IGF-1 działa na komórki tekalne, powodując zwiększoną produkcję androgenów. Komórki tekalne z kolei produkują czynnik martwicy nowotworu (TNF) i czynnik wzrostu naskórka (EGF), które są również regulowane przez FSH.
EGF stymuluje proliferację komórek ziarnistych. IGF-2 jest głównym czynnikiem wzrostu w płynie pęcherzykowym, a IGF-1, TNF-a, TNF-3 i EGF również się w nim znajdują.
Zaburzenie parakrynnej i/lub autokrynnej regulacji funkcji jajników wydaje się odgrywać rolę w zaburzeniach owulacji i rozwoju policystycznych jajników.
W miarę wzrostu pęcherzyka antralnego wzrasta zawartość estrogenu w płynie pęcherzykowym. W szczytowym momencie wzrostu receptory LH pojawiają się na komórkach ziarnistych, następuje luteinizacja komórek ziarnistych i wzrasta produkcja progesteronu. Tak więc w okresie przedowulacyjnym wzrost produkcji estrogenu powoduje pojawienie się receptorów LH, LH z kolei powoduje luteinizację komórek ziarnistych i produkcję progesteronu. Wzrost progesteronu obniża poziom estrogenu, co najwyraźniej powoduje drugi szczyt FSH w środku cyklu.
Uważa się, że owulacja następuje 10-12 godzin po szczycie LH i 24-36 godzin po szczycie estradiolu. Uważa się, że LH stymuluje podział redukcyjny oocytu, luteinizację komórek ziarnistych i syntezę progesteronu i prostaglandyn w pęcherzyku.
Progesteron zwiększa aktywność enzymów proteolitycznych, które razem z prostaglandyną uczestniczą w pękaniu ściany pęcherzyka. Indukowany progesteronem szczyt FSH pozwala oocytowi opuścić pęcherzyk poprzez przekształcenie plazminogenu w enzym proteolityczny plazminę i zapewnia wystarczającą liczbę receptorów LH do prawidłowego rozwoju fazy lutealnej.
W ciągu 3 dni po owulacji komórki ziarniste zwiększają swoją wielkość, a w nich pojawiają się charakterystyczne wakuole wypełnione pigmentem, luteiną. Komórki teka-lutealne różnicują się z teki i podścieliska i stają się częścią ciałka żółtego. Naczynia włosowate penetrujące ciałko żółte rozwijają się bardzo szybko pod wpływem czynników angiogenezy, a wraz z poprawą waskularyzacji wzrasta produkcja progesteronu i estrogenów. Aktywność steroidogenezy i długość życia ciałka żółtego są determinowane przez poziom LH. Ciałko żółte nie jest jednorodną formacją komórkową. Oprócz 2 typów komórek lutealnych zawiera komórki śródbłonka, makrofagi, fibroblasty itp. Duże komórki lutealne wytwarzają peptydy (relaksynę, oksytocynę) i są bardziej aktywne w steroidogenezie z większą aktywnością aromatazy i większą syntezą progesteronu niż małe komórki.
Szczyt progesteronu występuje 8. dnia po szczycie LH. Należy zauważyć, że progesteron i estradiol są wydzielane epizodycznie w fazie lutealnej w korelacji z pulsacyjnym wydzielaniem LH. Wraz z tworzeniem się ciałka żółtego kontrola produkcji inhibiny przechodzi z FSH na LH. Inhibina wzrasta wraz ze wzrostem estradiolu przed szczytem LH i nadal wzrasta po szczycie LH, chociaż poziom estrogenu spada. Chociaż inhibina i estradiol są wydzielane przez komórki ziarniste, są regulowane przez różne ścieżki. Spadek inhibiny pod koniec fazy lutealnej przyczynia się do wzrostu FSH w następnym cyklu.
Ciałko żółte zanika bardzo szybko - 9.-11. dnia po owulacji.
Mechanizm degeneracji jest niejasny i nie jest związany z luteolityczną rolą estrogenów ani z mechanizmem związanym z receptorami, jak zaobserwowano w endometrium. Istnieje inne wyjaśnienie roli estrogenów wytwarzanych przez ciałko żółte. Wiadomo, że estrogeny są niezbędne do syntezy receptorów progesteronu w endometrium. Estrogeny fazy lutealnej są prawdopodobnie niezbędne do zmian związanych z progesteronem w endometrium po owulacji. Niewystarczający rozwój receptorów progesteronu, w wyniku niewystarczającego poziomu estrogenu, jest prawdopodobnie dodatkowym mechanizmem niepłodności i wczesnych poronień, innej formy niedoboru fazy lutealnej. Uważa się, że długość życia ciałka żółtego ustala się w momencie owulacji. I z pewnością ulegnie regresji, jeśli nie będzie wspierana przez ludzką gonadotropinę kosmówkową z powodu ciąży. Tak więc regresja ciałka żółtego prowadzi do spadku poziomu estradiolu, progesteronu i inhibiny. Zmniejszenie inhibiny usuwa jej supresyjny wpływ na FSH; zmniejszenie estradiolu i progesteronu pozwala na bardzo szybkie przywrócenie wydzielania GnRH i usunięcie mechanizmu sprzężenia zwrotnego z przysadki mózgowej. Zmniejszenie inhibiny i estradiolu, wraz ze wzrostem GnRH, powoduje wzrost FSH w stosunku do LH. Wzrost FSH prowadzi do wzrostu pęcherzyka z późniejszym wyborem dominującego pęcherzyka, a nowy cykl rozpoczyna się, jeśli ciąża nie nastąpi. Hormony steroidowe odgrywają wiodącą rolę w biologii rozrodu i ogólnej fizjologii. Określają fenotyp człowieka, wpływają na układ sercowo-naczyniowy, metabolizm kości, skórę, ogólne samopoczucie i odgrywają kluczową rolę w ciąży. Działanie hormonów steroidowych odzwierciedla mechanizmy wewnątrzkomórkowe i genetyczne, które są niezbędne do przekazania sygnału zewnątrzkomórkowego do jądra komórkowego w celu wywołania reakcji fizjologicznej.
Estrogeny dyfundują przez błonę komórkową i wiążą się z receptorami zlokalizowanymi w jądrze komórkowym. Kompleks receptor-steroid wiąże się następnie z DNA. W komórkach docelowych interakcje te prowadzą do ekspresji genów, syntezy białek i specyficznej funkcji komórek i tkanek.