Ekspert medyczny artykułu

Nowe publikacje
przysadka mózgowa
Ostatnia recenzja: 04.07.2025

Cała zawartość iLive jest sprawdzana medycznie lub sprawdzana pod względem faktycznym, aby zapewnić jak największą dokładność faktyczną.
Mamy ścisłe wytyczne dotyczące pozyskiwania i tylko linki do renomowanych serwisów medialnych, akademickich instytucji badawczych i, o ile to możliwe, recenzowanych badań medycznych. Zauważ, że liczby w nawiasach ([1], [2] itd.) Są linkami do tych badań, które można kliknąć.
Jeśli uważasz, że któraś z naszych treści jest niedokładna, nieaktualna lub w inny sposób wątpliwa, wybierz ją i naciśnij Ctrl + Enter.
Przysadka mózgowa (hypophysis, s.glandula pituitaria) znajduje się w dole przysadkowym siodła tureckiego kości klinowej i jest oddzielona od jamy czaszki wyrostkiem opony twardej mózgu, tworząc przeponę siodła. Przez otwór w tej przeponie przysadka mózgowa łączy się z lejkiem podwzgórza międzymózgowia. Rozmiar poprzeczny przysadki wynosi 10-17 mm, przednio-tylny 5-15 mm, pionowy 5-10 mm. Masa przysadki mózgowej u mężczyzn wynosi około 0,5 g, u kobiet - 0,6 g. Przysadka mózgowa jest pokryta zewnętrznie torebką.
Zgodnie z rozwojem przysadki mózgowej z dwóch różnych zawiązków, w narządzie rozróżnia się dwa płaty - przedni i tylny. Adenohypophysis, czyli płat przedni (adenohypophysis, s.lobus anterior), jest większy i stanowi 70-80% całkowitej masy przysadki mózgowej. Jest gęstszy od płata tylnego. W płacie przednim wyróżnia się część dystalną (pars distalis), która zajmuje przednią część dołu przysadki, część pośrednią (pars intermedia), położoną na granicy z płatem tylnym, oraz część guzowatą (pars tuberalis), biegnącą ku górze i łączącą się z lejkiem podwzgórza. Ze względu na obfitość naczyń krwionośnych płat przedni ma jasnożółty kolor z czerwonawym odcieniem. Miąższ przedniego płata przysadki mózgowej jest reprezentowany przez kilka typów komórek gruczołowych, pomiędzy których pasmami znajdują się sinusoidalne naczynia włosowate. Połowa (50%) komórek przysadki mózgowej to adenocyty chromofilowe, które mają drobnoziarniste granulki w cytoplazmie, dobrze barwiące się solami chromu. Są to adenocyty acidofilne (40% wszystkich komórek przysadki mózgowej) i adenocyty zasadochłonne (10%). Adenocyty zasadochłonne obejmują endokrynocyty gonadotropowe, kortykotropowe i tyreotropowe. Adenocyty chromofobowe są małe, mają duże jądro i niewielką ilość cytoplazmy. Komórki te są uważane za prekursory adenocytów chromofilowych. Pozostałe 50% komórek przysadki mózgowej to adenocyty chromofobowe.
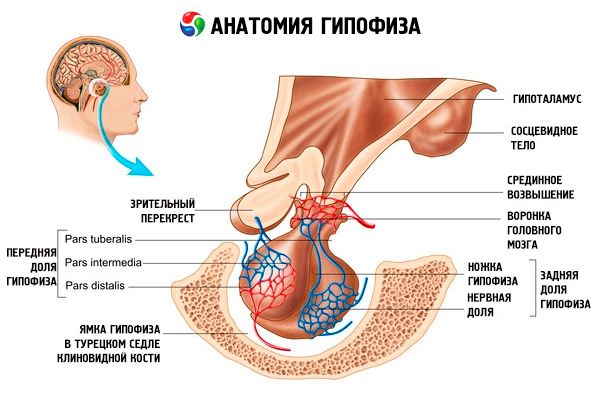
Neurohypophysis, czyli tylny płat (neurohypophysis, s.lobus posterior), składa się z płata nerwowego (lobus nervosus), który znajduje się w tylnej części dołu przysadki mózgowej, oraz lejka (infundibulum), znajdującego się za częścią guzowatą przysadki mózgowej. Tylny płat przysadki mózgowej tworzą komórki glejowe (komórki przysadki), włókna nerwowe, które biegną od jąder neurosekrecyjnych podwzgórza do neurohypophysis, oraz ciałka neurosekrecyjne.
Przysadka mózgowa, za pośrednictwem włókien nerwowych (ścieżek) i naczyń krwionośnych, jest funkcjonalnie połączona z podwzgórzem międzymózgowia, które reguluje aktywność przysadki mózgowej. Przysadka mózgowa i podwzgórze, wraz z ich połączeniami neuroendokrynnymi, naczyniowymi i nerwowymi, są zwykle uważane za układ podwzgórzowo-przysadkowy.
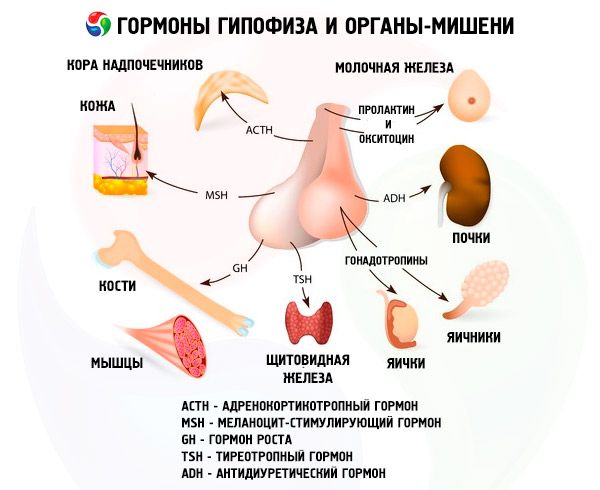
Hormony przedniego i tylnego płata przysadki mózgowej wpływają na wiele funkcji organizmu, przede wszystkim poprzez inne gruczoły dokrewne. W przednim płacie przysadki mózgowej, acidofilne adenocyty (komórki alfa) produkują hormon somatotropowy (hormon wzrostu), który uczestniczy w regulacji procesów wzrostu i rozwoju młodych organizmów. Endokrynocyty kortykotropowe wydzielają hormon adrenokortykotropowy (ACTH), który stymuluje wydzielanie hormonów steroidowych przez nadnercza. Endokrynocyty tyreotropowe wydzielają hormon tyreotropowy (TSH), który wpływa na rozwój tarczycy i aktywuje produkcję jej hormonów. Hormony gonadotropowe: folikulotropowy (FSH), luteinizujący (LH) i prolaktyna - wpływają na dojrzewanie płciowe organizmu, regulują i stymulują rozwój pęcherzyków w jajniku, owulację, wzrost gruczołów piersiowych i produkcję mleka u kobiet, proces spermatogenezy u mężczyzn. Hormony te są wytwarzane przez bazofilowe adenocyty (komórki beta). Wydzielane są tu również czynniki lipotropowe przysadki mózgowej, które wpływają na mobilizację i wykorzystanie tłuszczów w organizmie. W części pośredniej przedniego płata powstaje hormon stymulujący melanocyty, który kontroluje powstawanie pigmentów - melanin - w organizmie.
Komórki neurosekrecyjne jąder nadwzrokowego i przykomorowego podwzgórza produkują wazopresynę i oksytocynę. Hormony te są transportowane do komórek tylnego płata przysadki mózgowej wzdłuż aksonów tworzących układ podwzgórzowo-przysadkowy. Z tylnego płata przysadki mózgowej substancje te przedostają się do krwi. Hormon wazopresyna ma działanie zwężające naczynia krwionośne i antydiuretyczne, dlatego nazywany jest również hormonem antydiuretycznym (ADH). Oksytocyna działa pobudzająco na kurczliwość mięśni macicy, zwiększa wydzielanie mleka przez laktujący gruczoł piersiowy, hamuje rozwój i funkcję ciałka żółtego oraz wpływa na zmiany napięcia mięśni gładkich (nieprążkowanych) przewodu pokarmowego.
Rozwój przysadki mózgowej
Przedni płat przysadki mózgowej rozwija się z nabłonka ściany grzbietowej zatoki ustnej w postaci pierścieniowatego wyrostka (worek Rathkego). Ten ektodermalny występ rośnie w kierunku dna przyszłej trzeciej komory. W jego kierunku, od dolnej powierzchni drugiego pęcherzyka mózgowego (przyszłego dna trzeciej komory), rozwija się wyrostek, z którego rozwija się szary guzek lejka i tylny płat przysadki mózgowej.
Naczynia i nerwy przysadki mózgowej
Z tętnic szyjnych wewnętrznych i naczyń koła tętniczego mózgowia tętnice przysadkowe górne i dolne są kierowane do przysadki mózgowej. Tętnice przysadkowe górne trafiają do jądra szarego i lejka podwzgórza, tutaj zespalają się ze sobą i tworzą naczynia włosowate penetrujące tkankę mózgową - pierwotną sieć naczyń włosowatych. Z długich i krótkich pętli tej sieci powstają żyły wrotne, które są kierowane do przedniego płata przysadki mózgowej. W miąższu przedniego płata przysadki żyły te rozpadają się na szerokie sinusoidalne naczynia włosowate, tworząc wtórną sieć naczyń włosowatych. Tylny płat przysadki mózgowej jest zaopatrywany w krew głównie przez tętnicę przysadkową dolną. Pomiędzy tętnicami przysadkowymi górnymi i dolnymi występują długie zespolenia tętnicze. Odpływ krwi żylnej z wtórnej sieci naczyń włosowatych odbywa się poprzez układ żył, które wpływają do zatok jamistych i międzyjamistych opony twardej mózgu.
Włókna współczulne, które wchodzą do narządu wraz z tętnicami, uczestniczą w unerwieniu przysadki mózgowej. Postganglionowe włókna współczulne odchodzą od splotu tętnicy szyjnej wewnętrznej. Ponadto w tylnym płacie przysadki mózgowej znajdują się liczne zakończenia wypustek komórek neurosekrecyjnych zlokalizowanych w jądrach podwzgórza.
Cechy przysadki mózgowej związane z wiekiem
Średnia masa przysadki mózgowej u noworodków sięga 0,12 g. Masa narządu podwaja się do 10 roku życia, a potraja do 15 roku życia. W wieku 20 lat masa przysadki mózgowej osiąga maksimum (530-560 mg) i pozostaje prawie niezmieniona w kolejnych okresach wieku. Po 60 roku życia obserwuje się niewielki spadek masy tego gruczołu dokrewnego.
 [ 7 ], [ 8 ], [ 9 ], [ 10 ], [ 11 ], [ 12 ], [ 13 ], [ 14 ], [ 15 ]
[ 7 ], [ 8 ], [ 9 ], [ 10 ], [ 11 ], [ 12 ], [ 13 ], [ 14 ], [ 15 ]
Hormony przysadki mózgowej
Jedność regulacji nerwowej i hormonalnej w organizmie jest zapewniona przez ścisłe połączenie anatomiczne i czynnościowe przysadki mózgowej i podwzgórza. Kompleks ten determinuje stan i funkcjonowanie całego układu endokrynnego.
Głównym gruczołem dokrewnym, który produkuje szereg hormonów peptydowych, które bezpośrednio regulują funkcję gruczołów obwodowych, jest przysadka mózgowa. Jest to czerwonoszara fasolowata twór pokryty włóknistą torebką o wadze 0,5-0,6 g. W zależności od płci i wieku człowieka zmienia się nieznacznie. Nadal obowiązuje ogólnie przyjęty podział przysadki na dwa płaty różniące się rozwojem, budową i funkcją: przedni dystalny - przysadkę mózgową i tylny - przysadkę mózgową. Pierwszy stanowi około 70% całkowitej masy gruczołu i jest konwencjonalnie dzielony na część dystalną, lejkowatą i pośrednią, drugi - na część tylną, czyli płat, i szypułę przysadki. Gruczoł znajduje się w dole przysadkowym siodła tureckiego kości klinowej i jest połączony z mózgiem za pomocą szypuły. Górna część przedniego płata jest pokryta skrzyżowaniem wzrokowym i traktami wzrokowymi. Ukrwienie przysadki jest bardzo obfite i odbywa się za pomocą gałęzi tętnicy szyjnej wewnętrznej (tętnice przysadkowe górne i dolne), a także gałęzi koła tętniczego mózgu. Tętnice przysadkowe górne uczestniczą w ukrwieniu przysadki gruczołowej, a dolne – przysadki nerwowej, kontaktując się z zakończeniami neurosekrecyjnymi aksonów jąder wielkokomórkowych podwzgórza. Te pierwsze wchodzą do wyniosłości pośrodkowej podwzgórza, gdzie rozchodzą się w sieć naczyń włosowatych (splot naczyń włosowatych pierwotnych). Te naczynia włosowate (z którymi stykają się zakończenia aksonów małych komórek neurosekrecyjnych podwzgórza mediobasalnego) zbierają się w żyłach wrotnych schodzących wzdłuż szypuły przysadki do miąższu przysadki mózgowej, gdzie ponownie dzielą się na sieć sinusoidalnych naczyń włosowatych (splot naczyń włosowatych wtórnych). W ten sposób krew, po wcześniejszym przejściu przez wyniosłość środkową podwzgórza, gdzie zostaje wzbogacona o podwzgórzowe hormony adenohypofizotropowe (hormony uwalniające), wchodzi do przysadki mózgowej.
Odpływ krwi nasyconej hormonami przysadki mózgowej z licznych naczyń włosowatych splotu wtórnego odbywa się poprzez układ żył, które z kolei wpływają do zatok żylnych opony twardej, a następnie do ogólnego krwiobiegu. W ten sposób układ wrotny przysadki mózgowej ze zstępującym kierunkiem przepływu krwi z podwzgórza jest morfofunkcjonalnym składnikiem złożonego mechanizmu neurohumoralnej kontroli funkcji zwrotnikowych przysadki mózgowej.
Przysadka mózgowa jest unerwiona przez włókna współczulne, które podążają za tętnicami przysadkowymi. Mają swój początek w włóknach postganglionowych, które przechodzą przez splot tętnicy szyjnej wewnętrznej, połączonych z górnymi zwojami szyjnymi. Nie ma bezpośredniego unerwienia przysadki mózgowej z podwzgórza. Płat tylny otrzymuje włókna nerwowe z jąder neurosekrecyjnych podwzgórza.
Przysadka mózgowa jest bardzo złożoną formacją w swojej architekturze histologicznej. Posiada dwa rodzaje komórek gruczołowych - chromofobowe i chromofilowe. Te ostatnie dzielą się z kolei na acidofilne i zasadochłonne (szczegółowy opis histologiczny przysadki mózgowej znajduje się w odpowiedniej sekcji podręcznika). Należy jednak zauważyć, że hormony wytwarzane przez komórki gruczołowe tworzące miąższ przysadki mózgowej, ze względu na różnorodność tych ostatnich, są w pewnym stopniu różne pod względem natury chemicznej, a drobna struktura komórek wydzielniczych musi odpowiadać cechom biosyntezy każdej z nich. Czasami jednak w przysadce mózgowej można zaobserwować formy przejściowe komórek gruczołowych, które są zdolne do wytwarzania kilku hormonów. Istnieją dowody na to, że rodzaj komórek gruczołowych przysadki mózgowej nie zawsze jest uwarunkowany genetycznie.
Pod przeponą siodła tureckiego znajduje się lejkowata część płata przedniego. Obejmuje ona trzon przysadki mózgowej, stykając się z guzkiem szarym. Ta część przysadki mózgowej charakteryzuje się obecnością komórek nabłonkowych i obfitym ukrwieniem. Jest również aktywna hormonalnie.
Środkowa część przysadki mózgowej składa się z kilku warstw dużych, wydzielniczych, zasadochłonnych komórek.
Przysadka mózgowa wykonuje różne funkcje poprzez swoje hormony. Jej przedni płat produkuje hormony adrenokortykotropowe (ACTH), tyreotropowe (TSH), folikulotropowe (FSH), luteinizujące (LH), lipotropowe, a także hormon wzrostu - somatotropowy (STO) i prolaktynę. W płacie pośrednim syntetyzowany jest hormon melanocytotropowy (MSH), a w płacie tylnym kumuluje się wazopresyna i oksytocyna.
ACTH
Hormony przysadkowe to grupa hormonów białkowych i peptydowych oraz glikoprotein. Spośród hormonów przedniego płata przysadki mózgowej najlepiej zbadano ACTH. Jest on produkowany przez komórki zasadochłonne. Jego główną funkcją fizjologiczną jest stymulacja biosyntezy i wydzielania hormonów steroidowych przez korę nadnerczy. ACTH wykazuje również działanie pobudzające melanocyty i lipotropowe. W 1953 roku został wyizolowany w czystej postaci. Później ustalono jego strukturę chemiczną, składającą się z 39 reszt aminokwasowych u ludzi i wielu ssaków. ACTH nie ma specyficzności gatunkowej. Obecnie przeprowadzono syntezę chemiczną zarówno samego hormonu, jak i różnych fragmentów jego cząsteczki, bardziej aktywnych niż hormony naturalne. Struktura hormonu ma dwie sekcje łańcucha peptydowego, z których jedna zapewnia wykrywanie i wiązanie ACTH z receptorem, a druga zapewnia działanie biologiczne. Najwyraźniej wiąże się z receptorem ACTH poprzez interakcję ładunków elektrycznych hormonu i receptora. Rolę biologicznego efektora ACTH pełni fragment cząsteczki 4-10 (Met-Glu-His-Phen-Arg-Tri-Tri).
Aktywność ACTH stymulująca melanocyty wynika z obecności w cząsteczce regionu N-końcowego, składającego się z 13 reszt aminokwasowych i powtarzającego strukturę hormonu alfa-stymulującego melanocyty. Region ten zawiera również heptapeptyd obecny w innych hormonach przysadki mózgowej i posiadający pewne działania adrenokortykotropowe, stymulujące melanocyty i lipotropowe.
Za kluczowy moment w działaniu ACTH uważa się aktywację enzymu kinazy białkowej w cytoplazmie przy udziale cAMP. Fosforylowana kinaza białkowa aktywuje enzym esterazę, która przekształca estry cholesterolu w wolną substancję w kropelkach tłuszczu. Białko syntetyzowane w cytoplazmie w wyniku fosforylacji rybosomów stymuluje wiązanie wolnego cholesterolu do cytochromu P-450 i jego transfer z kropelek lipidowych do mitochondriów, gdzie obecne są wszystkie enzymy zapewniające przekształcenie cholesterolu w kortykosteroidy.
[ 16 ], [ 17 ], [ 18 ], [ 19 ], [ 20 ], [ 21 ], [ 22 ]
Hormon tyreotropowy
TSH - tyreotropina - główny regulator rozwoju i funkcjonowania tarczycy, procesów syntezy i wydzielania hormonów tarczycy. To złożone białko - glikoproteina - składa się z podjednostek alfa i beta. Struktura pierwszej podjednostki pokrywa się z podjednostką alfa hormonu luteinizującego. Ponadto w dużej mierze pokrywa się u różnych gatunków zwierząt. Sekwencja reszt aminokwasowych w podjednostce beta ludzkiego TSH została rozszyfrowana i składa się ze 119 reszt aminokwasowych. Można zauważyć, że podjednostki beta ludzkiego i bydlęcego TSH są pod wieloma względami podobne. Właściwości biologiczne i charakter aktywności biologicznej hormonów glikoproteinowych są determinowane przez podjednostkę beta. Zapewnia ona również interakcję hormonu z receptorami w różnych narządach docelowych. Jednak podjednostka beta u większości zwierząt wykazuje specyficzną aktywność dopiero po połączeniu z podjednostką alfa, która działa jako swoisty aktywator hormonu. Ten ostatni z równym prawdopodobieństwem wywołuje działanie luteinizujące, folikulotropowe i tyreotropowe, które są uwarunkowane właściwościami podjednostki beta. Stwierdzone podobieństwo pozwala nam wnioskować, że hormony te powstały w procesie ewolucji od jednego wspólnego poprzednika, podjednostka beta determinuje również właściwości immunologiczne hormonów. Istnieje przypuszczenie, że podjednostka alfa chroni podjednostkę beta przed działaniem enzymów proteolitycznych, a także ułatwia jej transport z przysadki mózgowej do obwodowych narządów docelowych.
Hormony gonadotropowe
Gonadotropiny występują w organizmie w postaci LH i FSH. Funkcjonalne przeznaczenie tych hormonów sprowadza się na ogół do zapewnienia procesów rozrodczych u osobników obu płci. Są one, podobnie jak TSH, złożonymi białkami - glikoproteinami. FSH indukuje dojrzewanie pęcherzyków w jajnikach u kobiet i stymuluje spermatogenezę u mężczyzn. LH powoduje pęknięcie pęcherzyka u kobiet z utworzeniem ciałka żółtego i stymuluje wydzielanie estrogenów i progesteronu. U mężczyzn ten sam hormon przyspiesza rozwój tkanki śródmiąższowej i wydzielanie androgenów. Efekty gonadotropin są od siebie zależne i występują synchronicznie.
Dynamika wydzielania gonadotropin u kobiet zmienia się w trakcie cyklu menstruacyjnego i została zbadana wystarczająco szczegółowo. W fazie przedowulacyjnej (folikularnej) cyklu zawartość LH jest na dość niskim poziomie, podczas gdy FSH jest zwiększone. W miarę dojrzewania pęcherzyka zwiększa się wydzielanie estradiolu, co sprzyja zwiększonej produkcji gonadotropin przez przysadkę mózgową i występowaniu cykli LH i FSH, tj. sterydy płciowe stymulują wydzielanie gonadotropin.
Obecnie ustalono strukturę LH. Podobnie jak TSH, składa się z 2 podjednostek: a i b. Struktura podjednostki alfa LH u różnych gatunków zwierząt w dużej mierze pokrywa się, odpowiada strukturze podjednostki alfa TSH.
Struktura podjednostki beta LH różni się znacząco od struktury podjednostki beta TSH, chociaż ma cztery identyczne odcinki łańcucha peptydowego składające się z 4-5 reszt aminokwasowych. W TSH zlokalizowane są one w pozycjach 27-31, 51-54, 65-68 i 78-83. Ponieważ podjednostka beta LH i TSH determinuje specyficzną aktywność biologiczną hormonów, można założyć, że homologiczne odcinki w strukturze LH i TSH powinny zapewniać połączenie podjednostek beta z podjednostką alfa, a odcinki różniące się budową powinny odpowiadać za specyficzność aktywności biologicznej hormonów.
Natywny LH jest bardzo odporny na działanie enzymów proteolitycznych, ale podjednostka beta jest szybko rozszczepiana przez chymotrypsynę, a podjednostka a jest trudna do zhydrolizowania przez enzym, tzn. pełni rolę ochronną, uniemożliwiając chymotrypsynie dostęp do wiązań peptydowych.
Jeśli chodzi o strukturę chemiczną FSH, badacze nie uzyskali jeszcze ostatecznych wyników. Podobnie jak LH, FSH składa się z dwóch podjednostek, ale podjednostka beta FSH różni się od podjednostki beta LH.
Prolaktyna
Inny hormon, prolaktyna (hormon laktogenny), odgrywa aktywną rolę w procesach rozrodczych. Główne właściwości fizjologiczne prolaktyny u ssaków objawiają się w postaci stymulacji rozwoju gruczołów mlekowych i laktacji, wzrostu gruczołów łojowych i narządów wewnętrznych. Promuje ona manifestację działania steroidów na drugorzędne cechy płciowe u samców, stymuluje aktywność wydzielniczą ciałka żółtego u myszy i szczurów oraz uczestniczy w regulacji metabolizmu tłuszczów. W ostatnich latach wiele uwagi poświęcono prolaktynie jako regulatorowi zachowań macierzyńskich; taką polifunkcyjność tłumaczy się jej ewolucyjnym rozwojem. Jest to jeden ze starożytnych hormonów przysadki mózgowej i występuje nawet u płazów. Obecnie struktura prolaktyny u niektórych gatunków ssaków została całkowicie rozszyfrowana. Jednak do niedawna naukowcy wyrażali wątpliwości co do istnienia takiego hormonu u ludzi. Wielu uważało, że jego funkcję pełni hormon wzrostu. Teraz uzyskano przekonujące dowody na obecność prolaktyny u ludzi, a jej struktura została częściowo rozszyfrowana. Receptory prolaktyny aktywnie wiążą hormon wzrostu i laktogen łożyskowy, co wskazuje na pojedynczy mechanizm działania trzech hormonów.
Somatotropina
Hormon wzrostu, somatotropina, ma jeszcze szersze spektrum działania niż prolaktyna. Podobnie jak prolaktyna, jest wytwarzany przez komórki acidofilne przysadki mózgowej. STH stymuluje wzrost szkieletu, aktywuje biosyntezę białek, ma działanie mobilizujące tłuszcz i promuje wzrost masy ciała. Ponadto koordynuje procesy metaboliczne.
Udział hormonu w tym ostatnim potwierdza fakt gwałtownego wzrostu jego wydzielania przez przysadkę mózgową, np. w przypadku obniżenia się poziomu cukru we krwi.
Struktura chemiczna tego ludzkiego hormonu jest obecnie w pełni ustalona - 191 reszt aminokwasowych. Jego struktura podstawowa jest podobna do struktury somatomammotropiny kosmówkowej lub laktogenu łożyskowego. Dane te wskazują na znaczną ewolucyjną bliskość obu hormonów, chociaż wykazują różnice w aktywności biologicznej.
Należy podkreślić wysoką specyficzność gatunkową omawianego hormonu – na przykład STH pochodzenia zwierzęcego jest nieaktywny u ludzi. Wyjaśnia to zarówno reakcja między receptorami STH u ludzi i zwierząt, jak i struktura samego hormonu. Obecnie trwają badania mające na celu identyfikację centrów aktywnych w złożonej strukturze STH, które wykazują aktywność biologiczną. Badane są poszczególne fragmenty cząsteczki, które wykazują inne właściwości. Na przykład po hydrolizie ludzkiego STH pepsyną wyizolowano peptyd składający się z 14 reszt aminokwasowych i odpowiadający sekcji cząsteczki 31-44. Nie miał on wpływu na wzrost, ale znacznie przewyższał natywny hormon pod względem aktywności lipotropowej. Ludzki hormon wzrostu, w przeciwieństwie do podobnego hormonu zwierzęcego, ma znaczną aktywność laktogenną.
Przysadka mózgowa syntetyzuje wiele substancji peptydowych i białkowych, które mają działanie mobilizujące tłuszcz, a hormony tropowe przysadki - ACTH, STH, TSH i inne - mają działanie lipotropowe. W ostatnich latach szczególnie podkreślono hormony beta- i y-lipotropowe (LPG). Właściwości biologiczne beta-LPG zostały zbadane najbardziej szczegółowo; oprócz aktywności lipotropowej ma również działanie stymulujące melanocyty, stymulujące kortykotropinę i hipokalcemiczne, a także wywołuje działanie podobne do insuliny.
Obecnie rozszyfrowano strukturę pierwotną LPG owczego (90 reszt aminokwasowych), lipotropowych hormonów świń i bydła. Hormon ten ma specyficzność gatunkową, chociaż struktura centralnego regionu beta-LPG jest taka sama u różnych gatunków. Określa ona właściwości biologiczne hormonu. Jeden z fragmentów tego regionu znajduje się w strukturze alfa-MSH, beta-MSH, ACTH i beta-LPG. Sugeruje się, że hormony te powstały z tego samego prekursora w procesie ewolucji. γ-LPG ma słabszą aktywność lipotropową niż beta-LPG.
Hormon stymulujący melanocyty
Ten hormon, syntetyzowany w pośrednim płacie przysadki mózgowej, stymuluje biosyntezę pigmentu skóry melaniny w jej funkcji biologicznej, promuje wzrost wielkości i liczby komórek pigmentowych melanocytów w skórze płazów. Te cechy MSH są wykorzystywane w biologicznych testach hormonu. Istnieją dwa rodzaje hormonu: alfa- i beta-MSH. Wykazano, że alfa-MSH nie ma specyficzności gatunkowej i ma taką samą strukturę chemiczną u wszystkich ssaków. Jego cząsteczka jest łańcuchem peptydowym składającym się z 13 reszt aminokwasowych. Beta-MSH natomiast ma specyficzność gatunkową, a jego struktura różni się u różnych zwierząt. U większości ssaków cząsteczka beta-MSH składa się z 18 reszt aminokwasowych, a tylko u ludzi jest wydłużona od końca aminokwasowego o cztery reszty aminokwasowe. Należy zauważyć, że alfa-MSH ma pewną aktywność adrenokortykotropową, a jego wpływ na zachowanie zwierząt i ludzi został już udowodniony.
Oksytocyna i wazopresyna
Wazopresyna i oksytocyna, które są syntetyzowane w podwzgórzu, gromadzą się w tylnym płacie przysadki mózgowej: wazopresyna w neuronach jądra nadoczodołowego, a oksytocyna w jądrze przykomorowym. Następnie są one przenoszone do przysadki mózgowej. Należy podkreślić, że prekursor hormonu wazopresyny jest syntetyzowany w podwzgórzu jako pierwszy. Jednocześnie produkowane są tam białka neurofizyny typu 1 i 2. Pierwsze wiąże oksytocynę, a drugie wazopresynę. Kompleksy te migrują w postaci granulek neurosekrecyjnych w cytoplazmie wzdłuż aksonu i docierają do tylnego płata przysadki mózgowej, gdzie włókna nerwowe kończą się w ścianie naczyniowej, a zawartość granulek przedostaje się do krwi. Wazopresyna i oksytocyna są pierwszymi hormonami przysadkowymi o całkowicie ustalonej sekwencji aminokwasowej. Pod względem struktury chemicznej są to nonapeptydy z jednym mostkiem disiarczkowym.
Rozważane hormony wywołują różne efekty biologiczne: stymulują transport wody i soli przez błony, działają wazopresyjnie, wzmacniają skurcze mięśni gładkich macicy podczas porodu i zwiększają wydzielanie gruczołów sutkowych. Należy zauważyć, że wazopresyna ma wyższą aktywność antydiuretyczną niż oksytocyna, podczas gdy ta druga ma silniejszy wpływ na macicę i gruczoł sutkowy. Głównym regulatorem wydzielania wazopresyny jest spożycie wody; w kanalikach nerkowych wiąże się ona z receptorami w błonach cytoplazmatycznych, a następnie aktywuje w nich enzym cyklazę adenylową. Różne części cząsteczki są odpowiedzialne za wiązanie hormonu z receptorem i za efekt biologiczny.
Przysadka mózgowa, połączona poprzez podwzgórze z całym układem nerwowym, jednoczy układ hormonalny w funkcjonalną całość, uczestnicząc w zapewnieniu stałości środowiska wewnętrznego organizmu (homeostazy). W obrębie układu hormonalnego regulacja homeostatyczna odbywa się na zasadzie sprzężenia zwrotnego między przednim płatem przysadki mózgowej a gruczołami „docelowymi” (tarczyca, kora nadnerczy, gonady). Nadmiar hormonu produkowanego przez gruczoł „docelowy” hamuje, a jego niedobór stymuluje wydzielanie i uwalnianie odpowiedniego hormonu tropowego. Podwzgórze jest włączone do układu sprzężenia zwrotnego. To w nim znajdują się strefy receptorowe wrażliwe na hormony gruczołów „docelowych”. Poprzez specyficzne wiązanie się z hormonami krążącymi we krwi i zmieniając odpowiedź w zależności od stężenia hormonów, receptory podwzgórzowe przekazują swój efekt do odpowiednich ośrodków podwzgórzowych, które koordynują pracę przysadki mózgowej, uwalniając podwzgórzowe hormony adenohypofizotropowe. Zatem podwzgórze należy uważać za mózg neuroendokrynny.